What Do Baby Snails Look Like When They Hatch Giant African Land Snail
Introduction
Invertebrates include approximately one million animal species kept both in zoological institutions and in households where they are hand raised equally exotic pets (1). Gastropods comprise nearly sixty,000 species of aquatic as well every bit terrestrial mollusks, primarily snails and slugs (1). The giant African land snail (Achatina fulica, syn. Lissachatina fulica) is native to East Africa, however it is a widespread invasive species in Asia, Oceania, and more recently in Americas, where it has been introduced accidentally or purposefully as a food source and as a pet. Its release in natural ecosystems, agricultural, and urban areas has resulted in ecological, health, and agricultural threats (2, 3). The giant African country snail is an intermediate hosts for several parasites including Aelurostrongylus abstrusus, Angiostrongylus cantonensis, Angiostrongylus costaricensis, Schistosoma mansoni, Hymenolepis spp., and Fasciola hepatica (4, 5). All the above helminths, with the exception of A. abstrusus, are able to cause severe diseases in humans. Specifically, the giant African land snail is the master gastropod responsible for worldwide spread of A. cantonensis that causes human eosinophilic meningoencephalitis in Asia and Americas (ii, 6). Take chances factors for infection in humans, pets and wild animals with those helminth parasites include the ingestion of raw or undercooked infected snails or slugs, or foods contaminated by the slime of infected snails or slugs (four, 5, vii, 8).
Although the African giant land snails are amidst the most popular snails held every bit pets, and their worldwide popularity as exotic pets is growing rapidly, only a few surveys have been carried out on the occurrence of their parasites in natural atmospheric condition (2, 6, 9–11). The aim of this survey was to investigate the occurrence of parasites in behemothic African land snails bred as pets in southern Italy.
Materials and Methods
General Data
In August 2018, three pools of fresh fecal samples obtained from a total of 60 giant African land snails, kept in 3 different private collections located in Pozzuoli, Caserta, and Naples (Italy), were investigated for parasites. From each locality we obtained a puddle of feces from twenty individual snails each. The snails included in this survey had a median age of i.half dozen years ranging from 0.2 to 2 years and had not received whatever previous anti-parasitic treatment. Snails were fed fresh vegetables and fruits; additionally, a calcium powder supplementation (Calcium, Exo Terra, Hagen Germany GmbH & Co. KG, Holm, Germany) was provided twice a week. All animals were bred in Italy, privately owned and kept as pets in 4 indoor terrariums of threescore × xxx × 45 cm in size, in groups of 10–20 animals (2 groups of 20 snails in Pozzuoli and Caserta and two groups of ten snails each in Naples). An organic peat soil (Organic Coco-Peat Soil, Eastward-Coco Products, Gloucestershire, UK), previously heat-treated (100°C for 30 min) and so free of parasites and insects, was used as substrate.
Diagnostic Procedures
Xx grams of fresh feces were initially obtained from each of the four terrarium. When pooled carrion analyzed by fresh smear, flotation and Baermann examination showed positivity to rhabditid nematodes, ii snails from each positive terrarium were housed individually in a sterile plastic box, and their carrion were collected, immediately subsequently defecation, in a sterile 50 ml plastic tube. Coprological examination for pooled and individual samples included fresh smear, centrifugal flotation (2 g of feces for each test) using a solution of sugar and formaldehyde (specific gravity 1.27) and Baermann test (10 1000 of feces for each test). Additionally, the mucus obtained during fresh smear test was analyzed nether a light microscope. Because the larval forms of several helminth species may encyst inside the foot muscle of snails, histological test of biopsy samples harvested from the anterior and posterior areas of the foot muscle was performed in 30 snails, anesthetized with the technique described by Giannelli et al. (12) and Gilbertson and Wyatt (thirteen). The biopsy samples were fixed in 10% neutral phosphate-buffered formalin, and candy by routine methods into paraffin blocks that were cutting into iii μm thick sections, and stained with hematoxylin and eosin. The study was performed under permission of the do where it took place and under signed, written informed owner consent.
Morphological Identification
Temporary mounts were made by heat-killing nematodes on glass slides in a driblet of water, after which a drinking glass embrace slip was applied. Nematodes were transferred into a drop of tap water on a glass slide and placed on heater (100°C) for 10 s. These specimens were used for morphological identification. An AMPLIVAL light microscope, Carl Zeiss Jena, and a Leitz Diaplan with Nomarski optics were used for observation. Morphological identification of rhabditid nematodes followed Andrassy (xiv) and Andrassy (15) and information technology was mostly based on the morphology of pharynx and stoma, and reproductive arrangement, mainly the features of spicules, number and position of papillae or presence and size or shape of bursa for males; and tail shape, position of vulva and morphology of reproductive arrangement for females. The identification of larvae was based on the tail shape.
Molecular Analysis
DNA was extracted from nematode specimens obtained from feces fixed in 96% ethanol nerveless from the three localities. Nematodes were washed in double-distilled water (ddHtwoO) overnight, prior to the molecular analysis, for complete removal of residuals of ethanol. Each individual nematode was transferred into a sterile Eppendorf tube (200 μl) with 20 μl of extraction buffer (17.7 μl of ddH2O, 2 μl of 10 × concentrated PCR buffer, 0.ii μl of 1% Tween xx and 0.1 μl of proteinase Yard). Buffer and nematodes were frozen at −20°C for 20 min, and then immediately incubated at 65°C for 1 h, followed past 10 min at 95°C. The lysates were cooled on ice and so centrifuged (2 min, ix,000 chiliad); 1 μl of supernatant was used for PCR.
A fragment of rDNA containing the internal transcribed spacer regions (ITS1, v.8S, ITS2) was amplified using primers 18S: 5′-TTG ATT ACG TCC CTG CCC TTT-3′ (forrad), and 28S: 5′-TTT CAC TCG CCG TTA CTA AGG-three′ (opposite) (xvi). A fragment of rDNA containing the factor for 18S rRNA was amplified using primers 22F: 5′- TCC AAG GAA GGC AGC AG GC-three′ (forward), and 1080JR: 5′- TCC TGG TGG TGC CCT TCC GTC AAT TTC-3′ (reverse) (17). The PCR master mix consisted of: ddHiiO, 7.25μl; 10 × PCR buffer, 1.25 μl; deoxynucleoside triphosphates (dNTPs), 1 μl; 0.75 μl of both forrad and contrary primer; polymerase, 0.ane μl; and 1 μl of Deoxyribonucleic acid-extract. The PCR profiles were used equally follows: for ITS 1 cycle of 94°C for 7 min followed by 35 cycles of 94°C for sixty s, l°C for 60 southward and 72°C for 60 s, and a final elongation at 72°C for 7 min (14); for 18S i cycle of 94°C for 5 min, followed by 35 cycles of 94°C for lx s, 55°C for 90 due south and 72°C for 2 min and a last elongation at 72°C for 10 min. The PCR products were sequenced by GATC Biotech (Frg) and later edited and uploaded to GenBank (https://www.ncbi.nlm.nih.gov/genbank/).
Results
The Baermann examination revealed the presence of nematode larvae, whose length ranged from 170 to 336 μm, in all samples. Eggs, larvae and developed rhabditid nematodes were detected by fresh smear and flotation methods in all fecal samples. Rhabditella axei (Figure one) was identified morphologically from two out of iii pooled and private fecal samples from Pozzuoli and Naples. In add-on, Rhabditis terricola, Cruznema sp., and Pristionchus entomophagus were isolated from ane pool and individual fecal samples from Caserta. No parasites were detected in the musculus biopsy samples or in the mucus.
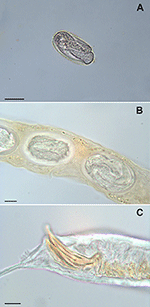
Figure one. Rhabditella axei obtained from the pet giant African country snails (Achatina fulica). An egg from coprological test, bar = xxx μm (A) eggs in uterus, bar = 10 μm (B) caudal extremity of male, bar = 10 μm (C).
Several morphological types of rhabditid nematodes were isolated from snail feces fixed in 96% ethanol. Obtained and edited sequences of ITS and 18S of these nematodes were compared (Blast) with the material in GenBank and identified every bit R. axei (fractional 18S, accretion number MK124578, similarity 99%), Rh. terricola (partial 18S and fractional ITS1, accession number MK156052, similarity 100%), P. entomophagus (fractional 18S and partial ITS1, accretion number MK156050, similarity 99%) and Cruznema sp. (fractional 18S and partial ITS1, accession number MK156051, similarity 96–100%).
Discussion
The results of this written report indicate that all the examined behemothic African land snails lay eggs, larvae, and developed rhabditid nematodes in the feces, and may therefore correspond a source of infection for other pets and humans. In club to avoid misidentification with Strongyloides sp., a nematode species showing close resemblance with rhabditids that has a clear parasitic zoonotic relevance (fourteen, xv, xviii), the initial morphological diagnosis of rhabditid nematodes was confirmed through molecular analyses.
Rhabditidae include free-living saprophytic nematodes, widely found in soil and organic debris where they feed mainly on bacteria. Many species of snails may serve as final definitive hosts for rhabditid nematodes (14, 15). However, a number of Rhabditis and Rhabditella species has been associated with vertebrates including humans (14, 15, nineteen–27). Although their presence can be the result of environmental contamination, these nematodes may cause diseases in many animals and humans. Rhabditis elongata, Rh. hominis, and Rh. usuii larvae have been isolated from human carrion, urine and vaginal swabs (19–21). Nevertheless, non many cases of symptomatic infections have been reported in humans (21–24). Feng and Li (25) described 2 human being cases of urinary infection by R. axei in China, and Ahn et al. (xx) reported 5 human cases of intestinal infection with Rhabditis sp. in rural schoolhouse children of South korea. Similarly, two cases of human intestinal infection by R. axei were described in China (26), whereas another published work (21) reported a case of intestinal infection in a v-month-onetime Brazilian child who was presented with fever and watery and bloody diarrhea; coprological examination revealed eggs, larvae and adults of Rhabditis sp. Meamar et al. (27) described the occurrence of watery diarrhea in ii Iranian patients with AIDS, associated to astringent intestinal infection past larvae and adult specimens of R. axei. Finally, Teschner et al. (24) recently described a case of outer ear canal infection in a 37 twelvemonth-old human presented with purulent otorrhea from both ears and acute hearing loss caused past Rhabditis sp. In full general, Rhabditis spp. are considered a common crusade of external otitis in cattle living in tropical areas (e.g., south America and Africa), peculiarly in older animals, and accept been identified likewise in chickens, dogs and pigs with incoercible diarrhea (18, 28–30). Even so, asymptomatic infections accept also been described and these nematodes are frequently considered pseudoparasites (28, 29).
Although free-living nematodes were retrieved in all fecal samples, no specimens were constitute in the mucus and histological samples. A possible explanation for this finding is that nematodes may locate in unlike tissues/organs of their hosts, depending on the type of association nematode/mollusk host (xv, 31, 32). Rhabditis spp. complete its life bicycle within the snail, with no damage to its molluscan host. Previous studies on the costless-ranging African snails (Archachatina spp. and Achatina spp.) revealed that R. axei lives in the gastrointestinal tract of its snail host where the entire nematode life cycle is completed (31–33).
Although African giant snails are listed as hosts for R. axei, the occurrence of this nematode had been reported only in a few species of giant African snails other than A. fulica, namely Archachatina marginata ovum, Ar. marginata saturalis, and Achatina achatina (31). On the other hand, P. entomophagus has a worldwide distribution and has been mainly associated to dung beetles belonging to the superfamily Scarabaeoidea (34, 35), R. terricola has been constitute in salamanders (xiv, fifteen) and Cruznema spp. in the cricket Gryllodes laplatae (Orthoptera) (36). In all cases, manual of the parasite occurs through the contact of the snails with contaminated moist soil that is rich in decomposing organic matter (31, 37). Although, in the present cases all the animals were kept in terraria with a heat-treated organic soil, we speculate that infection may accept occurred before buy of the animals at the pet shops or breeding facilities.
In conclusion, our results signal that the pet behemothic African land snails may serve equally reservoir of several rhabditid nematodes. This snail species is amid the most commonly ones kept as pets, and therefore often live in shut proximity to humans. As a event, the contamination of the domestic environment through their feces is possible. Although parasitic nematodes were not isolated in this study, the giant African country snails should still be considered potential carriers of nematodes able to crusade opportunistic diseases in humans. Therefore, we highlight the importance of further epidemiological enquiry on the occurrence of costless-living and parasitic nematodes in gastropod snails kept in captivity, and emphasize the need for strict control measures to reduce the take chances of opportunistic infection with rhabditid nematodes in pet snail owners.
Author Contributions
Dd'O and MS conceived and planned the analysis. MS and JN carried out the assay. CA helped shape the inquiry and edited the manuscript. All authors provided critical feedback and contributed to the final manuscript.
Disharmonize of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of involvement.
References
i. Zachariah T, Mitchell M. Invertebrates. In: Mitchell MA, Tully T, editors. Manual of Exotic Pet Practice. St. Louis, MO: Saunders Elsevier (2009). p. eleven–38.
Google Scholar
ii. Ohlweiler FP, de Almeida Guimarães MC, Takahashi FY, Eduardo JM. Current distribution of Achatina fulica, in the land of São Paulo including records of Aelurostrongylus abstrusus (nematoda) larvae infestation. Rev Inst Med Trop Sao Paulo. (2010) 52:211–14. doi: x.1590/S0036-46652010000400009
PubMed Abstract | CrossRef Full Text | Google Scholar
three. Raut SK, Baker GM. Achatina fulica Bowdich and other Achatinidae as pests in tropical agriculture. In: Baker GM, editor. Molluscs as Crop Pests. Wallingford: CAB International (2002). p. 55–114.
5. Igbinosa IB, Isaac C, Ojagefu Adamu H, Adeleke G. Parasites of edible land snails in Edo State, Nigeria. Helminthologia. (2016) 53:331–5. doi: 10.1515/helmin-2016-0031
CrossRef Full Text | Google Scholar
6. Vitta A, Polseela R, Nateeworanart S, Tattiyapong M. Survey of Angiostrongylus cantonensis in rat and giant African land snails in Phitsanulok province. Asian Pac J Trop Med. (2011) 4:597–9. doi: ten.1016/S1995-7645(eleven)60154-5
CrossRef Full Text | Google Scholar
seven. Alfaro-Alarcón A, Veneziano 5, Galiero G, Cerrone A, Gutierrez Northward, Chinchilla A, et al. Offset study of a naturally patent infection of Angiostrongylus costaricensis in a dog. Vet Parasitol. (2015) 212:431–four. doi: ten.1016/j.vetpar.2015.08.016
PubMed Abstruse | CrossRef Total Text | Google Scholar
8. Santoro M, Alfaro-Alarcón A, Veneziano V, Cerrone A, Latrofa MS, Otranto D, et al. The white-nosed coati (Nasua narica) is a naturally susceptible definitive host for the zoonotic nematode Angiostrongylus costaricensis in Costa rica. Vet. Parasitol. (2016) 228:93–5. doi: ten.1016/j.vetpar.2016.08.017
PubMed Abstract | CrossRef Full Text | Google Scholar
9. Liboria M, Morales G, Carmen S, Isbelia S, Luz AP. First finding in Venezuela of Schistosoma mansoni eggs and other helminths of involvement in public health plant in faeces and mucous secretion of the mollusc Achatina fulica (Bowdich, 1822). Zootecnia Trop. (2010) 28:83–394.
Google Scholar
x. Moreira VLC, Giese EG, Melo FTV, Simoes RO, Thiengo SC, Maldonado A, et al. Endemic angiostrongyliasis in the Brazilian Amazon: natural parasitism of Agiostrongylus cantonensis in Rattus rattus and R. norvegicus, and sympatric giant African land snail, Achatina fulica. Acta Tropica. (2013) 125:90–7. doi: x.1016/j.actatropica.2012.x.001
CrossRef Full Text | Google Scholar
11. Iwanowicz DD, Sanders LR, Schill WB, Xayavong MV, da Silva AJ, Qvarnstrom Y, et al. Spread of the rat lungworm (Angiostrongylus cantonensis) in giant African land snails (Lissachatina fulica) in FLorida, The states. J Wildl Dis. (2015) 51:749–53. doi: 10.7589/2014-06-160
PubMed Abstract | CrossRef Total Text | Google Scholar
12. Giannelli A, Colella V, Abramo F, do Nascimento Ramos RA, Falsone 50, Brianti East, et al. Release of lungworm larvae from snails in the environment: potential for alternative manual pathways. PLoS Negl Trop Dis. (2015) 9:e0003722. doi: x.1371/journal.pntd.0003722
PubMed Abstruse | CrossRef Full Text | Google Scholar
13. Gilbertson CR, Wyatt JD. Evaluation of euthanasia techniques for an invertebrate species, land snails (Succinea putris). J Am Assoc Lab Anim Sci. (2016) 55:577–81.
PubMed Abstract | Google Scholar
14. Andrassy I. A Taxonomic Review of the Suborder Rhabditina (Nematoda: Secernentia). Paris: ORSTOM (1983). p. 241.
Google Scholar
15. Andrassy I. Klasse Nematoda. Berlin: Akademie Verlag (1984). p. 512.
16. Vrain TC, Wakarchuk DA, Levesque Air conditioning, Hamilton RI. Intraspecific rDNA restriction fragment length polymorphism in the Xiphinema americanum group. Fundam Appl Nematol. (1992) 15:563–73.
Google Scholar
17. Ross JL, Ivanova ES, Spiridonov SE, Waeyenberge 50, Moens M, Nicol GW, et al. Molecular phylogeny of slug-parasitic nematodes inferred from 18S rRNA gene sequences. Mol Phylogenet Evol. (2010) 55:738–43. doi: x.1016/j.ympev.2010.01.026
PubMed Abstract | CrossRef Total Text | Google Scholar
18. Stachurska-Hagen T, Johnsen OH, Robertson LJ. Not-Strongyloides rhabditida identified in fecal samples – two case reports: lessons learned from morphological and molecular diagnostic approaches. Parasitol Open. (2016) 2:1–8. doi: 10.1017/pao.2016.11
CrossRef Full Text | Google Scholar
xix. Faust EC, Rusell PF, Jung RC. Craig and Faust's Clinical Parasitology. London: Kimpton (1970). p. 890.
Google Scholar
twenty. Ahn YK, Chung PR, Lee KT. Rhabditis sp. infected cases in rural school children. Kisaengchunghak Chapchi. (1985) 23:i–six.
PubMed Abstract | Google Scholar
21. Campos DMB, Araújo JL, Vieira MCM, Damasceno F, Barbosa AP. A case of parasitism by Rhabditis sp. in a kid from Goiânia, Goiás, Brazil. Rev Soc Bras Med Trop. (2002) 35:519–22. doi: x.1590/S0037-86822002000500016
PubMed Abstruse | CrossRef Full Text | Google Scholar
22. Eldridge B. Human urinary tract infection acquired by Rhabditis sp. PHLS Microbiol Digest. (1993) 10:202–3.
23. He YX, Jiang H. Three human cases of urinary tract infection with Rhabditis. Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi. (1985) 3:206–viii.
Google Scholar
24. Teschner K, Würfel W, Sedlacek L, Suerbaum South, Tappe D, Hornef MW. Outer ear canal infection with Rhabditis sp. nematodes in a human. J Clin Microbiol. (2014) 52:1793–five.
PubMed Abstract | Google Scholar
25. Feng LC, Li F. 2 human being cases of urinary tract infection with Rhabditella axei. Peking Nat Hist Bull. (1950) xviii:195–202.
Google Scholar
26. Ye LP, Zhu CG, Zhang JN. Two cases of Rhaditis axei infections in human being digestive organisation. Chin J Schistosomiasis Control. (2002) 14:187.
27. Meamar AR, Kia EB, Zahabiun F, Jafari-Mehr A, Moghadam A, Sadjjadi SM. The occurrence of severe infections with Rhabditis axei in AIDS patients in Islamic republic of iran. J Helminthol. (2007) 81:351–two. doi: x.1017/S0022149X07792301
PubMed Abstract | CrossRef Full Text | Google Scholar
28. Levine ND, Birch CL, Dolowy WC, McKinney RE. Rhabditis axei, a pseudoparasitic nematode of the dog. J Am Vet Med Assoc. (1963) xv:1404–half-dozen.
Google Scholar
29. El-Azazy O, El-Gawady H, Aught M. The occurrence of Rhabditis (Rhabditella) axei in the faeces of a chicken in Egypt. J Helminthol. (1988) 62:219–20.
PubMed Abstract | Google Scholar
30. Adewole Then, Fabumuy AO, Agunbiade RO, Ayeni SK. Occurrence of Rhabditis dubia in cattle dungs in Lagos State, Nigeria. J Environ Earth Sci. (2014) 4:26–viii.
Google Scholar
31. Odaibo AB, Dehinbo AJ, Olofintoye LK, Falode OA. Occurrence and distribution of Rhabditis axei in African behemothic snails in southwestern Nigeria. Helminthologia. (2000) 37:233–5.
Google Scholar
32. Grewal PS, Grewal SK, Tan 50, Adams BJ. Parasitism of molluscs by nematodes: types of associations and evolutionary trends. J Nematol. (2003) 35:146–56.
PubMed Abstract | Google Scholar
33. Olofintoye LK, Olorunniyi OF. Intensity of Rhabditis axei at different parts of gastrointestinal tracts of Archachatina marginata ovum and Achatina achatina. J Bio Innov. (2016) 5:182–5.
34. Herrmann Thousand, Mayer We, Sommer RJ. Nematodes of the genus Pristionchus are closely associated with scarab beetles and the Colorado spud beetle in Western Europe. Zoology. (2006) 109:96–108. doi: 10.1016/j.zool.2006.03.001
PubMed Abstract | CrossRef Full Text | Google Scholar
35. Kanzaki N, Ragsdale EJ, Herrmann Chiliad, Sommer RJ. Two new and two recharacterized species from a radiation of Pristionchus. (Nematoda: Diplogastridae) in Europe. J Nematol. (2014) 46:threescore–74.
PubMed Abstruse | Google Scholar
36. Rebored GR, Camino NB. Two new species of nematodes (Rhabditida: Diplogasteridae and Rhabditidae) parasites of Gryllodes laplatae. (Orthoptera: Gryllidae) in Argentina. Mem Inst Oswaldo Cruz. (1998) 93:763–6.
Google Scholar
37. Nguyen KB. Methodology, morphology and identification. In: Nguyen KB, Hunt DJ, editors. Entomopathogenic Nematodes: Systematics, Phylogeny and Bacterial Symbionts. Nematology Monographs and Perspectives. Leiden: Brill (2007). p. 59–119.
What Do Baby Snails Look Like When They Hatch Giant African Land Snail
Source: https://www.frontiersin.org/articles/10.3389/fvets.2019.00088/full
0 Response to "What Do Baby Snails Look Like When They Hatch Giant African Land Snail"
Post a Comment